Peptidoglycan
{kind=link}
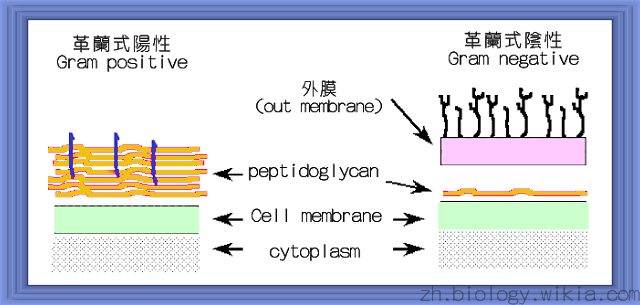
革蘭氏陽性菌和陰性菌細胞壁的比較
細菌的表面包有一層細胞壁,細胞壁的主要功能在於維持細菌形狀,並具有保護細菌的功能。此外,細菌的細胞壁也提供一個通透性障壁供特定分子的進出,同時也扮演著細胞與細胞間辨識及吸附的角色。細菌細胞壁所含的主要成分是peptidogylcan,gram positive bacteria和gram negative bacteria細胞壁的組成在結構上有所不同,可依組成結構的不同來分辨兩者,gram negative bacteria所含的 peptidoglycan layer較薄(只有單層),位於outer membrane 和 inner membrane兩層膜之間,然而gram positive bacteria所含的peptidoglycan layer較厚,並堆疊在細胞膜外。
Peptidogylcan是一種多醣的聚合物,其單體主要由N-acetylmuramic acid (NAM)和N-acetylglucosamine (NAG)兩個單醣所組成,兩者以β1→4 (O-link)的方式鍵結,可和其他單體利用相同方式交替相接成精密的長鏈,並再以短鏈的peptide互相串連形成三度空間的立體結構。這些連接用的短鏈peptide group通常是由五個amino acid所組成,與兩個單醣中的NAM相接,五個amino acid依序分別是L-Alanine, D-Glutamate, L-Lysine (或Diaminopimelic acid, DAP)和2 D-Alanine。然而gram positive bacteria和gram negative bacteria在peptide上的組成有所差異,gram positive bacteria短鏈peptide group中所含的第三個amiono acid是DAP,然而gram negative bacteria通常使用L-Lysine。
Gram positive bacteria和gram negative bacteria所含的peptidoglycan layer除了厚度以及連接用的peptide group組成上有所不同外,附著在peptidoglycan上的蛋白質也有所不同。在革蘭氏陽性菌中,其細胞壁中亦含有teichoic acid(台口酸) ,這個teichoic acid是由甘油或是含5個碳的酒精(叫做ribotol)所形成的多聚體(polymer),通常由5~30個單元所組成,本身會由膜中突出來而連接到peptidoglycan的骨架上。這個互相交錯所形成的共價結構,可被視為包圍在細胞外的一個巨大結構。一般說來,革蘭氏陰性菌的細胞壁有20~40層的peptidoglycan,與lipoprotein組合而成,且其厚度約為50 nm。
Peptidoglycan 的合成[ | ]
peptidoglycan的合成大致上可分為四個步驟。Peptidoglycan組合的第一步驟是先將sugar變成amino sugar,藉由UDP的活化,使UDP-GlcNAc利於進入Peptidoglycan的合成路徑。Amino sugar進一步變成UDP-NAM。新合成的UDP-NAM依序和三個amino acid組成的peptide連接,再接上D-Ala-D-Ala。NAM-peptide group和膜上的lipid carrier結合,再接上NAG,經由translocation到細胞膜外,再進一步進行polymerization。
Step 1. UDP-amino sugar的合成
- 形成amino sugar的過程是由fructose-6-P開始,經由Fructose-6-P amidotransferase和glutamine參與,生成glucosamine-6-P。接著Phosphoglucosamine mutase將glucosamine-6-P第六號位置上的磷酸根移到一號碳的位置上,形成glucosamine-1-P,這個步驟為可逆反應。Glucosamine-1-P再利用GlmU syntase使參與反應的acetyl-CoA將其acetyl group轉移到glucosamine-1-P第二個位置上,形成N-acetylglucosamine-1-P。GlmU syntase接著將UTP接到N-acetylglucosamine-1-P的磷酸根的位置上,並放出兩個磷酸根,形成UDP-NAG。
- PEP與UDP-NAG作用,接在第三號碳的OH基上,形成phospholactoyl-UDP-GlcNAc,經由MurA transferase作用,使PEP去掉一個磷酸根,形成EP-UDP-GlcNAc,接著NADPH和MurB reductase的參與,使EP-UDP-GlcNAc還原成UDP-NAM。
Step 2. UDP-NAM pentapeptide的合成
- 活化的UDP-NAM利用MurC、MurD、MurE和MurF依序接上L-Ala、D-Glu、NDP or L-Lys最後接上D-Ala-D-Ala。每一個步驟都會耗掉一個ATP。
- 然而自然界中的amino acid主要是以L form存在,因此細菌發展出一個特有的酵素—Racemase--可將L form的glutamate轉變成D-glutamate。另一個形成D-glutamate的方式是利用D-amino acid aminotransferase將D-Alanine上的胺基轉到α-keto-glutarate上形成D-glutamate。方程式如下:
D-Alanine +α-keto-glutarate ←→ D-gluamate + pyruvate
Step 3. Translocation
- 當Peptidoglycan的單元形成後,可接到膜上一個lipid carrier,這個lipid carrier的主要功能就是將接在carrier上的peptidoglycan單元,翻轉至細菌外,再作polymerization,主要有平面和立體的連接方式,平面的連接方式是NAG和NAM間的連接,立體的連接方式則是靠著tetrapeptide和tetrapeptide間的cross linkage。
- NAG和NAM間的連結主要靠著MurG (N-acertylglucosaminyl transferase),NAM-MAG兩兩相連後,釋出的free lipid carrier會進行去磷酸的反應,然後可翻轉回細胞膜內重覆使用。
Step 4. Polymerization and cross linking
- Tetrapeptide間的連結主要靠著Mur synthetase,peptide group間的連結是靠著一串glycine相連,一端與第三個amino aicd—L-lys相接,另一端則接在另一個tetrapeptide上第四個amino acid—D-Ala上,第五個D-Alanine被釋出。
應用|Application[ | ]
抗生素作用的機制不外乎抑制細菌與生理作用相關構造的合成,可分成下列四種:(1) 抑制細胞壁的合成;(2) 抑制蛋白質的合成;(3) 抑制核酸合成;(4) 改變細胞膜功能。本文僅就抑制細胞壁合成討論之。 能抑制細胞壁合成的抗生素有下列幾類:
- β-lactam antibiotics
- 這類抗生素都含有一個β-lactam ring的結構,常見的有penicillin and cephalosporins等等。
- Cycloserine:
- D-cycloserine結構類似於D-Alanine,它可和形成D-Alanine dipeptide和MurF的substrate binding site作用,防止D-Ala結合到NAM的peptide上。
- Fosfomycin (Phosphonomycin)
- 在合成的一開始,UDP-NAG變成UDP-NAM的步驟可被phosphnomycin抑制,使UDP-NAM無法合成,因此在Peptidoglycan的組成中便缺少了一個很重要的基本單元,細菌無法形成完整的Peptidoglycan,維持滲透壓的構造被破壞,便容易受外界影響而且打破。
- Vancomycin
- vancomycin是一種Transglycosylase抑制劑,可和peptidoglycan單元末端的D-Ala-D-Ala結合,形成五個氫鍵,同時抑制了transpeptidation and transgylcosylation。微生物的應變方式是將D-Ala-D-Ala改為D-Ala-D-lac或D-Ala-D-Ser。當D-Ala-D-Ala變成D-Ala-D-Ser時,氫鍵的作用位置改變或失去,使得兩者間的親合力下降了一千倍。(Walsh, C. Nature 2000)
- Ristocetin
- Transglycosylase抑制劑,可和新合成的D-Ala-D-Ala結合。
- Bacitracin
- Bacitracin是一種lipid phosphatase抑制劑,可干擾lipid carrier的作用,使lipid carrier交換NAM-NAG之後,無法翻轉回細胞內再利用。
|
|
|
| |
|
|
Penicillins |
|
|
|
| |||
|
Cycloserine |
|
| |
|
|
|
| |
|
|
|
|
|
|
Ristocetin |
|
| |
|
Bacitracin |
|
| |
參考資料|Reference[ | ]
- Eggert, U. S., N. Ruiz, et al. (2001). "Genetic basis for activity differences between vancomycin and glycolipid derivatives of vancomycin." Science 294(5541): 361-4.
- Goffin, C. and J. M. Ghuysen (1998). "Multimodular penicillin-binding proteins: an enigmatic family of orthologs and paralogs." Microbiol Mol Biol Rev 62(4): 1079-93.
- Iwaki, D., H. Mitsuzawa, et al. (2002). "The extracellular toll-like receptor 2 domain directly binds peptidoglycan derived from Staphylococcus aureus." J Biol Chem 277(27): 24315-20.
- Kengatharan Km Fau - De Kimpe, S., C. De Kimpe S Fau - Robson, et al. (1998). "Mechanism of gram-positive shock: identification of peptidoglycan and lipoteichoic acid moieties essential in the induction of nitric oxide synthase, shock, and multiple organ failure." J Exp Med 188(2): 305-15.
- Lazar, K. and S. Walker (2002). "Substrate analogues to study cell-wall biosynthesis and its inhibition." Curr Opin Chem Biol 6(6): 786-93.
- Merkel, G. J. and B. A. Scofield (2001). "Characterization of a monoclonal antibody that binds to an epitope on soluble bacterial peptidoglycan fragments." Clin Diagn Lab Immunol 8(3): 647-51.
- Ming, L. J. and J. D. Epperson (2002). "Metal binding and structure-activity relationship of the metalloantibiotic peptide bacitracin." J Inorg Biochem 91(1): 46-58.
- Murakami, S., D. Iwaki, et al. (2002). "Surfactant protein A inhibits peptidoglycan-induced tumor necrosis factor-alpha secretion in U937 cells and alveolar macrophages by direct interaction with toll-like receptor 2." J Biol Chem 277(9): 6830-7.
- Nanninga N. (1998). "Morphogenesis of Escherichia coli." Microbiol Mol Biol Rev 62(1):110-29
- Nau, R. and H. Eiffert (2002). "Modulation of release of proinflammatory bacterial compounds by antibacterials: potential impact on course of inflammation and outcome in sepsis and meningitis." Clin Microbiol Rev 15(1): 95-110.
- Navarre, W. W. and O. Schneewind (1999). "Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope." Microbiol Mol Biol Rev 63(1): 174-229.
- van Heijenoort, J. (1998). "Assembly of the monomer unit of bacterial peptidoglycan." Cell Mol Life Sci 54(4): 300-4.
- van Heijenoort, J. (2001). "Formation of the glycan chains in the synthesis of bacterial peptidoglycan." Glycobiology 11(3): 25R-36R.
- van Heijenoort, J. (2001). "Recent advances in the formation of the bacterial peptidoglycan monomer unit." Nat Prod Rep 18(5): 503-19.
- Walsh, C. (2000). "Molecular mechanisms that confer antibacterial drug resistance." Nature 406(6797): 775-81.
- Wiedemann, I., E. Breukink, et al. (2001). "Specific binding of nisin to the peptidoglycan precursor lipid II combines pore formation and inhibition of cell wall biosynthesis for potent antibiotic activity." J Biol Chem 276(3): 1772-9.
- http://www.ikepu.com.cn/datebase/details/scientist/19st/fleming.htm
討論[ | ]
|
|